Thylacocephala
 | |
|---|---|
 | |
| Classificazione scientifica | |
| Dominio | Eukaryota |
| Regno | Animalia |
| Sottoregno | Eumetazoa |
| Ramo | Bilateria |
| Superphylum | Protostomia |
| Phylum | Arthropoda |
| Classe | Thylacocephala Pinna et al., 1982 |
| Ordini | |
| |
Con il termine tilacocefali (Thylacocephala, dal greco θύλακος thylakos cioè "sacco" e κεφαλή kephalēs "testa") si indica una classe estinta di artropodi con carapace bivalve e addome ridotto non segmentato, di affinità incerta e tuttora in discussione, occorrenti dal Paleozoico inferiore al Cretacico.
Distribuzione stratigrafica
I fossili appartenenti a questo gruppo sono piuttosto rari, ma hanno una distribuzione stratigrafica ampia, che va dall'Ordoviciano superiore (Sandbiano)[1] al Cretacico superiore (Santoniano),[2]. È incerta (e oggetto di discussione) la loro presenza fin dal Cambriano.[3][4][5][6] La loro distribuzione geografica è quasi ubiquitaria: si rinvengono infatti in Australia, Giappone, Cina, Austria, Repubblica Ceca, Francia, Italia, Spagna, Scozia, Slovenia, Libano, Madagascar, Stati Uniti, Messico.[7]
Descrizione

La morfologia di questo gruppo è peculiare:[9][10] si tratta di artropodi a marcata simmetria bilaterale, caratterizzati da un carapace sottile e non mineralizzato a due valve identiche unite nella regione dorsale (prive di una struttura a cerniera definita), lateralmente compresso, di forma sub-ovale, ellittica o sub-trapezoidale, che racchiude quasi tutto il corpo ad eccezione dell'estrema parte posteriore. Addome ridotto non flessibile; appendici caudali e telson tipicamente assenti. Otto paia di branchie generalmente ben sviluppate. La conservazione del materiale fossile, anche per la presenza del carapace che avvolge le parti molli, non offre una visione esaustiva della segmentazione del corpo (numero e distribuzione dei somiti), che è tuttora materia di discussione tra gli esperti.
Il margine dorsale separa le due valve del carapace e la sua parte frontale reca una cresta che si prolunga in un rostro più o meno lungo e affilato. La parte anteriore del carapace è concava e mostra un paio di occhi composti globulari, sovente ipertrofici, che sporgono da un incavo situato tra il rostro frontale e il margine del carapace. Questi occhi nei fossili meglio conservati presentano un fitto pattern regolare con elevata densità di ommatidi, simile a quello di crostacei attuali.[11] Presenza di alcune (almeno 5) appendici cefaliche corte e scarsamente conosciute dal materiale fossile: probabilmente due paia di brevi antennule, tre paia di appendici mascellari e una sorta di labbro che formano l'apparato boccale dell'organismo.[12]

La nota più caratteristica[9][10][13] è la presenza di tre paia di lunghe appendici raptatorie,[14] prensili, genicolate[15] e dotate di spine, talora provviste di chele), che sporgono dal margine antero-ventrale del carapace, di cui il terzo paio particolarmente sviluppato. Il margine posteriore del carapace è concavo e beante, e da esso sporge la parte posteriore del corpo (quando conservata), che reca diverse (7-8, fino a 11) serie di segmenti pleurali rettangolari orientati perpendicolarmente al margine stesso, che recano appendici ventrali poco sviluppate, stiliformi e filamentose, con funzione probabilmente natatoria e/o deambulatoria, simili ai pleopodi presenti nella parte ventrale dell'addome dei crostacei, decrescenti in dimensioni verso la parte posteriore. I margini anteriore e/o posteriore possono essere dotati di spine; talvolta si ha un rostro posteriore. Le appendici, nei fossili meglio conservati, evidenziano la presenza di setae.[16]
In uno studio più recente, sono state riscontrate in talune forme ulteriori strutture enigmatiche[17]:
- una struttura sub-rettangolare allungata posteriore alla serie di segmenti pleurali, recante setae nella parte distale, che potrebbe essere interpretata come un "telson" ridotto (forse corrispondente alla parte interpretata come addome nella descrizione originaria di Ostenocaris cypriformis);[18]
- una "struttura circolare" concentrica situata sul "telson", interpretabile forse come ano oppure come statocisti;
- un paio di "strutture ovali" piatte situate subito dietro le appendici raptatorie, caratterizzate da una ampia superficie e margini recanti setae, probabilmente con funzioni natatorie o forse di ventilazione delle branchie.
Superficie esterna convessa; spesso si osserva una carena dorsale dove si uniscono le due valve del carapace e talora carene e solchi laterali longitudinali più o meno sviluppati. Ornamentazione generalmente non molto sviluppata, con striature, corrugamenti, serie lineari o concentriche di tubercoli o piccole depressioni (pori)[19], e talvolta sottili coste verticali sinuose.[20][21]
-
 Fossile di tilacocefalo (Dollocaris ingens) dal giacimento lagerstätte di La Voulte, Francia (Giurassico medio - Calloviano inferiore). Nel fossile, oltre al carapace sono visibili gli occhi, elementi del rostro e le appendici raptatorie. Parte anteriore verso sinistra.
Fossile di tilacocefalo (Dollocaris ingens) dal giacimento lagerstätte di La Voulte, Francia (Giurassico medio - Calloviano inferiore). Nel fossile, oltre al carapace sono visibili gli occhi, elementi del rostro e le appendici raptatorie. Parte anteriore verso sinistra. -
 Dollocaris ingens da La Voulte, Francia (Giurassico medio - Calloviano inferiore). Particolare delle appendici ("zampe" raptatorie). Parte anteriore verso sinistra. Notare che la chiusura delle zampe è dal basso verso l'alto.
Dollocaris ingens da La Voulte, Francia (Giurassico medio - Calloviano inferiore). Particolare delle appendici ("zampe" raptatorie). Parte anteriore verso sinistra. Notare che la chiusura delle zampe è dal basso verso l'alto. -
 Un crostaceo attuale, uno stomatopode (canocchia); notare le appendici anteriori raptatorie: si tratta di appendici specializzate per la predazione. Anche qui la chiusura è dal basso verso l'alto.
Un crostaceo attuale, uno stomatopode (canocchia); notare le appendici anteriori raptatorie: si tratta di appendici specializzate per la predazione. Anche qui la chiusura è dal basso verso l'alto. -
 Esemplare giovanile di Dollocaris ingens da La Voulte in nodulo fosfatico. Notare l'occhio ipertrofico e sporgente dall'incavo orbitale.
Esemplare giovanile di Dollocaris ingens da La Voulte in nodulo fosfatico. Notare l'occhio ipertrofico e sporgente dall'incavo orbitale. -
 Dollocaris ingens da La Voulte in nodulo fosfatico. Parte anteriore. Particolare di occhio.
Dollocaris ingens da La Voulte in nodulo fosfatico. Parte anteriore. Particolare di occhio. -
![Carapace di Ostenocaris cypriformis, Giurassico inferiore, conservato presso il Museo Stoppani di Venegono Inferiore.]]](//upload.wikimedia.org/wikipedia/commons/thumb/5/5c/Ostenocaris_cypriformis_455.JPG/244px-Ostenocaris_cypriformis_455.JPG) Carapace di Ostenocaris cypriformis, Giurassico inferiore, conservato presso il Museo Stoppani di Venegono Inferiore.]]
Carapace di Ostenocaris cypriformis, Giurassico inferiore, conservato presso il Museo Stoppani di Venegono Inferiore.]] -
 Carapace di Mayrocaris bucculata, del tardo Giurassico.
Carapace di Mayrocaris bucculata, del tardo Giurassico. -
 Esemplare di Clausocaris ribeti, tardo Giurassico. Notare l'enorme occhio e le appendici raptatorie estremamente sviluppate.
Esemplare di Clausocaris ribeti, tardo Giurassico. Notare l'enorme occhio e le appendici raptatorie estremamente sviluppate.





![Carapace di Ostenocaris cypriformis, Giurassico inferiore, conservato presso il Museo Stoppani di Venegono Inferiore.]]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/5c/Ostenocaris_cypriformis_455.JPG/244px-Ostenocaris_cypriformis_455.JPG)


Classificazione e caratteristiche morfologiche
I tilacocefali sono stati in origine riconosciuti come crostacei di un nuovo genere tra i fossili del giacimento sinemuriano di Osteno, sul Lago di Lugano. Inizialmente, queste forme vennero assegnate ad un nuovo genere di cirripedi (Ostenia cypriformis Arduini, Pinna, Teruzzi 1980)[22] Questa forma, ad un esame di maggiore dettaglio comprendente l'asportazione meccanica del carapace, considerate le peculiarità dei dettagli anatomici riconosciuti, venne più tardi assegnata ad una nuova classe di artropodi incertae sedis (Thylacocephala Pinna, Arduini, Pesarini, Teruzzi 1982),[23] della quale tuttavia veniva ipotizzata un'affinità filogenetica rispetto ai cirripedi.[24] Al momento dell'istituzione non venne data una definizione formale della classe, bensì sei caratteristiche diagnostiche:[25]
- porzione anteriore del cephalon enormemente sviluppata, modificata in forma di sacco, provvista di cuticola a scleriti, contenente il sacco ovarico; occhi assenti;
- carapace cefalico univalve racchiudente gran parte del cephalon e tutto il torace;
- antennule, antenne e massillipede molto sviluppati, uniramosi[26], modificati in appendici ambulatorie;
- apparato boccale composto di mandibole e maxille filtranti;
- torace ridotto rispetto al cephalon, composto da un primo segmento modificato in senso cefalico seguito da 8 somiti dotati, ciascuno, di una coppia di corti toracopodi;
- addome fortemente regredito e non segmentato.
Dopo questo primo lavoro pionieristico, varie osservazioni e controversie nell'ambito della comunità scientifica hanno portato a una revisione rispetto alla descrizione e all'interpretazione originaria;[27] tra i principali elementi di novità:
- la presenza di residui di organismi (pesci, uncini di cefalopodi, resti di crostacei) entro il sistema digestivo;[28]
- il riconoscimento di occhi composti di notevoli dimensioni (precedentemente interpretati come "sacco cefalico" privo di occhi), che hanno portato a correzioni sostanziali nell'interpretazione di questi organismi, non più come organismi sessili (solo occasionalmente e limitatamente deambulanti) bensì mobili e probabilmente predatori;[29]
- il "sacco ovarico" descritto originariamente (contenente elementi interpretati come uova)[30] si è rivelato in realtà un insieme di elementi vertebrali di pesci (quindi parte del contenuto dell'apparato digerente),[31] rendendo improbabile la presenza di strutture riproduttive nel segmento cefalico; questo implicherebbe anche la presenza di uno "stomaco" posto in posizione molto avanzata (tra gli occhi e poco dietro di questi) mentre l'"intestino" molto sviluppato ad anse descritto originariamente corrisponde in realtà alle strutture branchiali;[32]
Negli anni successivi, queste forme sono state revisionate e riconosciute in un gran numero di ambiti di età, orizzonti stratigrafici e condizioni diverse di fossilizzazione, portando al riconoscimento della maggior parte delle caratteristiche note di queste forme, tra le quali in particolare:[33]
- l'organizzazione bivalve del carapace;
- la funzione raptatoria delle appendici toraciche;
- la presenza di appendici posteriori con funzione deambulatoria o natatoria;
- la presenza (verificata per lo meno in alcune forme) di antennule e antenne ventralmente al rostro, elemento che le avvicina ai crostacei e che rende improbabile l'origine delle appendici raptatorie dalla modificazione di antenne.[34]
Il quadro della fisiologia e delle relazioni filogenetiche di queste creature è tuttavia ancora lontano da un completo chiarimento.[35]
-
![Ricostruzione di tilacocefalo Ostenocaris cypriformis secondo l'interpretazione originale risalente ai primi anni '80 del secolo scorso, accanto a un esemplare fossile.[36] Qui la parte anteriore dell'organismo viene ipotizzata come rivolta verso il basso e a contatto col sedimento (parzialmente infossata). Le appendici sono ricostruite come arti per la deambulazione.[37]](//upload.wikimedia.org/wikipedia/commons/thumb/a/af/Ostenocaris_cypriformis.JPG/215px-Ostenocaris_cypriformis.JPG) Ricostruzione di tilacocefalo Ostenocaris cypriformis secondo l'interpretazione originale risalente ai primi anni '80 del secolo scorso, accanto a un esemplare fossile.[36] Qui la parte anteriore dell'organismo viene ipotizzata come rivolta verso il basso e a contatto col sedimento (parzialmente infossata). Le appendici sono ricostruite come arti per la deambulazione.[37]
Ricostruzione di tilacocefalo Ostenocaris cypriformis secondo l'interpretazione originale risalente ai primi anni '80 del secolo scorso, accanto a un esemplare fossile.[36] Qui la parte anteriore dell'organismo viene ipotizzata come rivolta verso il basso e a contatto col sedimento (parzialmente infossata). Le appendici sono ricostruite come arti per la deambulazione.[37] -
![Ricostruzione di Ostenocaris cypriformis aggiornata rispetto agli studi successivi. La parte anteriore è rivolta verso l'alto e le appendici posteriori hanno funzione deambulatoria e/o natatoria[38], mentre le appendici più sviluppate sono rivolte in avanti, con funzione raptatoria.[39][40].](//upload.wikimedia.org/wikipedia/commons/thumb/0/0e/Ostenocaris_cypriformis_new.png/240px-Ostenocaris_cypriformis_new.png) Ricostruzione di Ostenocaris cypriformis aggiornata rispetto agli studi successivi. La parte anteriore è rivolta verso l'alto e le appendici posteriori hanno funzione deambulatoria e/o natatoria[38], mentre le appendici più sviluppate sono rivolte in avanti, con funzione raptatoria.[39][40].
Ricostruzione di Ostenocaris cypriformis aggiornata rispetto agli studi successivi. La parte anteriore è rivolta verso l'alto e le appendici posteriori hanno funzione deambulatoria e/o natatoria[38], mentre le appendici più sviluppate sono rivolte in avanti, con funzione raptatoria.[39][40]. -
![Modello di paleoarte di Kitakamicaris utatsuensis, un tilacocefalo del Triassico inferiore del Giappone. La ricostruzione artistica è corretta e basata su rinvenimenti fossili più recenti e completi (le appendici sono ipotetiche in quanto di queste forme sono stati rinvenuti i carapaci). In questo caso abbiamo un'ornamentazione sviluppata a coste sinuose.[41]](//upload.wikimedia.org/wikipedia/commons/thumb/5/5a/Kitakamicaris_utatsuensis_EcoSch_Tokyo2.jpeg/105px-Kitakamicaris_utatsuensis_EcoSch_Tokyo2.jpeg) Modello di paleoarte di Kitakamicaris utatsuensis, un tilacocefalo del Triassico inferiore del Giappone. La ricostruzione artistica è corretta e basata su rinvenimenti fossili più recenti e completi (le appendici sono ipotetiche in quanto di queste forme sono stati rinvenuti i carapaci). In questo caso abbiamo un'ornamentazione sviluppata a coste sinuose.[41]
Modello di paleoarte di Kitakamicaris utatsuensis, un tilacocefalo del Triassico inferiore del Giappone. La ricostruzione artistica è corretta e basata su rinvenimenti fossili più recenti e completi (le appendici sono ipotetiche in quanto di queste forme sono stati rinvenuti i carapaci). In questo caso abbiamo un'ornamentazione sviluppata a coste sinuose.[41] -
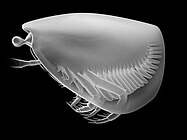 Thylacares brandonensis, un tilacocefalo del Siluriano.
Thylacares brandonensis, un tilacocefalo del Siluriano. -
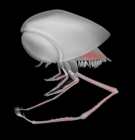 Clausocaris lithographica (Giurassico superiore), dal Calcare di Solnhofen.
Clausocaris lithographica (Giurassico superiore), dal Calcare di Solnhofen.
![Ricostruzione di tilacocefalo Ostenocaris cypriformis secondo l'interpretazione originale risalente ai primi anni '80 del secolo scorso, accanto a un esemplare fossile.[36] Qui la parte anteriore dell'organismo viene ipotizzata come rivolta verso il basso e a contatto col sedimento (parzialmente infossata). Le appendici sono ricostruite come arti per la deambulazione.[37]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/af/Ostenocaris_cypriformis.JPG/215px-Ostenocaris_cypriformis.JPG)
![Ricostruzione di Ostenocaris cypriformis aggiornata rispetto agli studi successivi. La parte anteriore è rivolta verso l'alto e le appendici posteriori hanno funzione deambulatoria e/o natatoria[38], mentre le appendici più sviluppate sono rivolte in avanti, con funzione raptatoria.[39][40].](http://upload.wikimedia.org/wikipedia/commons/thumb/0/0e/Ostenocaris_cypriformis_new.png/240px-Ostenocaris_cypriformis_new.png)
![Modello di paleoarte di Kitakamicaris utatsuensis, un tilacocefalo del Triassico inferiore del Giappone. La ricostruzione artistica è corretta e basata su rinvenimenti fossili più recenti e completi (le appendici sono ipotetiche in quanto di queste forme sono stati rinvenuti i carapaci). In questo caso abbiamo un'ornamentazione sviluppata a coste sinuose.[41]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/5a/Kitakamicaris_utatsuensis_EcoSch_Tokyo2.jpeg/105px-Kitakamicaris_utatsuensis_EcoSch_Tokyo2.jpeg)


Classificazione interna
La classe dei tilacocefali è suddivisibile in due ordini[42]:
- Concavicarida Briggs & Rolfe, 1983, costituita da tilacocefali con un carapace dotato di un apparato rostrale prominente che sovrasta anteriormente un incavo ottico ben definito;
- Conchyliocarida Secrétan, 1983, formata da tilacocefali dotati di un incavo visivo e un rostro mal definiti e di occhi situati sulla superficie di un "sacco" cefalico di grandi dimensioni.
Questa suddivisione pone l'accento sull'organizzazione dell'apparato visivo e sugli annessi elementi dell'esoscheletro ma non evidenzia differenze in altri elementi anatomici (appendici, segmentazione). [43]
Attualmente si riconoscono 24 generi:
- Ainiktozoon
- Ankitokazocaris
- Atropicaris
- Austriocaris
- Bohemiacaris
- Clausocaris
- Concavicaris
- Convexicaris
- Coreocaris
- Dollocaris
- Harrycaris
- Kilianicaris
- Kitakamicaris
- Mayrocaris
- Microcaris
- Ostenocaris
- Paraostenia
- Protozoea
- Pseuderichthus
- Pseodoprotozoea
- Thylacocephalus
- Rugocaris
- Yangzicaris
- Zhenghecaris
Stile di vita e habitat
Il modo di vita dei tilacocefali è stato a lungo dibattuto e rimane in gran parte una questione aperta. Inizialmente[44], erano interpretati come organismi sessili e filtratori, privi di occhi, parzialmente fossori[45] (il "sacco cefalico" era concepito come un vero e proprio organo fossatore), con affinità o convergenza evolutiva verso i cirripedi; questa interpretazione è stata in parte corretta successivamente[46] come organismi predatori, dotati di occhi composti (inizialmente interpretati come "sacco cefalico"), ma comunque essenzialmente bentonici e poco mobili; soprattutto con ridotte capacità di nuoto attivo, per la mancanza di un addome flessibile e di un telson e per le dimensioni ridotte delle appendici addominali. Osservazioni più recenti su materiale fossile meglio conservato però sembrano indicare che almeno alcune forme fossero capaci di nuoto attivo, per diverse caratteristiche[47]


- l'assenza di appendici toraciche sviluppate per la deambulazione, differentemente dai crostacei bentonici (le appendici raptatorie non sono adeguate in quanto specializzate per la predazione);
- il carapace relativamente sottile e non mineralizzato, quindi leggero;
- la presenza di numerose serie di appendici stiliformi, sporgenti dal margine postero-ventrale (pleopodi?), che suggeriscono una funzione locomotoria; la presenza di setae e di una muscolatura sviluppata potrebbe implicare una funzione nectonica.[49]
Lo stile di vita predatorio è indicato dalla presenza delle appendici raptatorie, molto robuste e provviste di spine, assai simili a quelle degli attuali stomatopodi (canocchie), che probabilmente venivano proiettate in avanti molto rapidamente chiudendosi dal basso verso l'alto sulla preda, che veniva in tal modo immobilizzata, forse in parte smembrata e portata all'apparato boccale. Una conferma viene dal contenuto dello stomaco di forme fossili (Ostenocaris) del Sinemuriano (Giurassico inferiore) lombardo[46] che consiste in resti di pesci, uncini di cefalopodi e frammenti di carapace di altri crostacei (inclusi tilacocefali di dimensioni minori). Queste caratteristiche si adattano peraltro sia a un carnivoro predatore che a un necrofago (in tal caso le appendici potevano servire per smembrare carcasse di organismi morti), o a entrambi gli adattamenti.[50]
Altri, osservando che le appendici raptatorie dei tilacocefali sono differenti come numero e disposizione dei segmenti rispetto a quelle dei crostacei attuali[51] e quindi un movimento simile a quello degli stomatopodi sarebbe stato forse meno efficiente, propendono per un diverso meccanismo di estensione: per lo meno in alcune forme, le appendici raptatorie non sarebbero state rivolte in avanti, in asse con il corpo, ma estese a 45° circa formando una sorta di "cesto" o di rete per intrappolare le prede.[52]
La mancanza di bioturbazione nei depositi fossiliferi e di organismi sicuramente fossori associati a queste forme[53][54] depone a favore di uno stile di vita non endobionte. La presenza in diverse forme di occhi composti molto sviluppati, simili a quelli di attuali forme di crostacei necto-planctonici (Hyperiidea) sembra indicare un adattamento a condizioni di elevata profondità del battente d'acqua e conseguente scarsa illuminazione; l'alta densità di ommatidi e implicherebbe una buona risoluzione visiva, con la capacità di distinguere piccoli oggetti.[55][56] La presenza sul carapace di talune forme di allineamenti di tubercoli o di depressioni fa peraltro pensare a possibili organi bioluminescenti, analogamente ad alcuni crostacei attuali di mare profondo, finalizzati ad attirare prede, o in funzione antipredatoria, o ancora di richiamo sessuale.[54][57]
L'habitat di elezione di queste forme, almeno nel Mesozoico, sembra essere di mare profondo, al di sotto dei 200 metri, rientrante quindi nel piano batiale, in un contesto quindi di scarpata continentale o di transizione scarpata-bacino, dominato da sedimenti fini fangosi.[58] Per le forme del Giurassico inferiore rinvenute nel bacino lombardo (Osteno - Calcare di Moltrasio), questa connotazione ambientale va declinata in un contesto locale di fondale emipelagico soggetto a episodi anossici,[53] nell'ambito di un bacino tettonico a prevalente sedimentazione torbiditica fine di materiale carbonatico, in un quadro geodinamico tettonico di rift.[59] I tilacocefali studiati nell'ambito dei famosi "fish beds" del Cretacico superiore del Libano (giacimento di Sahel Aalma)[60] sembrano ugualmente appartenere ad un contesto di mare piuttosto profondo (maggiore di 150 m), confermato dall'associazione faunistica.[61] Forme più antiche e problematiche (Cambriano inferiore) sono interpretate come predatori demersali di minore profondità (100-150 m, quindi nell'ambito della piattaforma continentale), in grado di nuotare liberamente ma legati al fondale per il nutrimento.[62] D'altro canto, alcune tra le prime forme attribuite con maggiore confidenza ai tilacocefali (Ordoviciano superiore) sono state rinvenute in un contesto di mare relativamente poco profondo (Letná Formation - Bacino Boemico) con sedimenti siltoso-arenacei ricchi di faune fossili ben differenziate, e sembrano mostrare in base alla morfologia del carapace una precoce differenziazione paleoecologica tra forme nectoniche (con carapace maggiormente "idrodinamico") e forme necto-bentoniche (con carapace massiccio e ovoidale) adattate ad ambiente con relativamente più alta energia del mezzo.[63]
-
 Ricostruzione artistica di una scena ipotetica relativa al giacimento di Osteno (Sinemuriano - Giurassico inferiore). Tilacocefali Ostenocaris cypriformis sono a caccia di cefalopodi (Ostenoteuthis) e di altri artropodi (Coleia, Phlyctisoma). Due si nutrono della carcassa di un conspecifico. Il sedimento, ricchissimo di spicole di spugne silicee, indica che queste forme erano comuni e formavano vere e proprie "praterie" sul fondale. Lo squaliforme Paleospinax pinnai passa sullo sfondo.
Ricostruzione artistica di una scena ipotetica relativa al giacimento di Osteno (Sinemuriano - Giurassico inferiore). Tilacocefali Ostenocaris cypriformis sono a caccia di cefalopodi (Ostenoteuthis) e di altri artropodi (Coleia, Phlyctisoma). Due si nutrono della carcassa di un conspecifico. Il sedimento, ricchissimo di spicole di spugne silicee, indica che queste forme erano comuni e formavano vere e proprie "praterie" sul fondale. Lo squaliforme Paleospinax pinnai passa sullo sfondo. -
![Una scena ricostruita ipoteticamente sulla base del materiale fossile del giacimento di La Voulte (Calloviano). Un tilacocefalo Dollocaris sp. tenta di predare un cefalopode di mare profondo (Proteroctopus ribeti); un altro Dollocaris inizia a smembrare una carcassa di pesce teleosteo per nutrirsene. Il fondale è ricoperto di spugne silicee; gli artropodi sono il gruppo faunistico dominante; nella colonna d'acqua sono presenti pesci e cefalopodi.[64]](//upload.wikimedia.org/wikipedia/commons/thumb/a/ab/Dollocaris_Proteroctopus_La_Voulte.png/445px-Dollocaris_Proteroctopus_La_Voulte.png) Una scena ricostruita ipoteticamente sulla base del materiale fossile del giacimento di La Voulte (Calloviano). Un tilacocefalo Dollocaris sp. tenta di predare un cefalopode di mare profondo (Proteroctopus ribeti); un altro Dollocaris inizia a smembrare una carcassa di pesce teleosteo per nutrirsene. Il fondale è ricoperto di spugne silicee; gli artropodi sono il gruppo faunistico dominante; nella colonna d'acqua sono presenti pesci e cefalopodi.[64]
Una scena ricostruita ipoteticamente sulla base del materiale fossile del giacimento di La Voulte (Calloviano). Un tilacocefalo Dollocaris sp. tenta di predare un cefalopode di mare profondo (Proteroctopus ribeti); un altro Dollocaris inizia a smembrare una carcassa di pesce teleosteo per nutrirsene. Il fondale è ricoperto di spugne silicee; gli artropodi sono il gruppo faunistico dominante; nella colonna d'acqua sono presenti pesci e cefalopodi.[64]

![Una scena ricostruita ipoteticamente sulla base del materiale fossile del giacimento di La Voulte (Calloviano). Un tilacocefalo Dollocaris sp. tenta di predare un cefalopode di mare profondo (Proteroctopus ribeti); un altro Dollocaris inizia a smembrare una carcassa di pesce teleosteo per nutrirsene. Il fondale è ricoperto di spugne silicee; gli artropodi sono il gruppo faunistico dominante; nella colonna d'acqua sono presenti pesci e cefalopodi.[64]](http://upload.wikimedia.org/wikipedia/commons/thumb/a/ab/Dollocaris_Proteroctopus_La_Voulte.png/445px-Dollocaris_Proteroctopus_La_Voulte.png)
Note
- ^ Van Roy et al. (2021).
- ^ Hady et al. (2024), capitolo: Sahel Aalma Lagerstätte.
- ^ Vannier et al. (2006)
- ^ Zeng et al. (2018)
- ^ Pates et al. (2021), pp. 7-8
- ^ L'estensione di questo gruppo al Cambriano inferiore si basa sulla discussa attribuzione sistematica della specie Zhenghecaris shankouensis Vannier et al. 2006, che viene dagli istitutori attribuita ai Thylacocephala, mentre successivamente è stata interpretata come parte del carapace di un artropode primitivo radiodonte (Zeng et al. 2018). Le argomentazioni da entrambe le parti non sono conclusive, e la posizione sistematica di questo taxon rimane incerta (Pates et al. 2021).
- ^ Ehiro et al. (2015), p. 269.
- ^ Vannier et al. (2006), p. 206; fig. 4, modificata.
- ^ a b Vannier et al. (2006), pp. 203-204; fig. 4.
- ^ a b Charbonnier et al. (2010), pp. 117-121.
- ^ Charbonnier et al. (2010), p. 119.
- ^ Vannier et al. (2006), p. 207.
- ^ Laville et al. (2023), pp. 10-50.
- ^ Appendici specializzate per la predazione, atte ad afferrare e trattenere la preda per portarla all'apparato boccale; un tipico esempio è il paio di zampe anteriori delle mantidi.
- ^ Con giunture a forma di ginocchio.
- ^ Haug et al. (2014), pp. 6, 7, 9, 13.
- ^ Laville et al. (2023), pp. 45-51; fig. 23.
- ^ Pinna et al. (1982), fig. 3.
- ^ Vannier et al. (2006), pp. 206-207.
- ^ Lange et al. (2001), fig. 1-4.
- ^ Ehiro et al. (2015), p. 272; fig.4.
- ^ Pinna et al. (1982), p. 470.
- ^ Pinna et al. (1982), pp. 476-480.
- ^ Pinna et al. (1982), p. 480.
- ^ Pinna et al. (1982), p. 476.
- ^ Sviluppati in un solo ramo, a differenza ad esempio, dei trilobiti.
- ^ Pinna et al. (1985).
- ^ Pinna et al. (1985), p. 373.
- ^ Pinna et al. (1985), p. 373.
- ^ Pinna et al. (1982), pp. 473.
- ^ Vannier et al. (2006), p. 210.
- ^ Van Roy et al. (2021), p. 2.
- ^ Vannier et al. (2006), p. 203-204, tab. 1
- ^ Lange et al. (2001), p. 907-911, fig. 1-4.
- ^ Vannier et al. (2006), p. 211.
- ^ Entrambi, fossile e modello, sono esposti nelle sale dl Museo civico di storia naturale di Milano.
- ^ Pinna et al. (1982), p. 474, 476-477; fig. 2-4.
- ^ Nei fossili di Osteno sono presenti solo le parti basali delle appendici, molto corte. Qui sono rappresentate ipoteticamente delle estensioni a spatola formate da setae che non sono conservate nel sito di Osteno, ma risultano presenti in altre forme da altri siti
- ^ Vannier et al. (2006).
- ^ Laville et al. (2023)
- ^ Ehiro et al. (2015).
- ^ Schram (1990).
- ^ Vannier et al. (2006).
- ^ Pinna et al. (1982), pp. 478-480.
- ^ Infossati nel sedimento, in parte endobionti
- ^ a b Pinna et al. (1985).
- ^ Charbonnier et al. (2010), pp.119-121.
- ^ Jobbins et al. (2020), Fig. 3
- ^ Haug et al. (2014), p. 13.
- ^ Vannier et al. (2006), p. 209.
- ^ Ad esempio, i segmenti di quelle della canocchia formano una "z", permettendone la chiusura completa anteriormente.
- ^ Haug et al. (2014), pp. 13-15.
- ^ a b Pinna (1985), p. 171.
- ^ a b Charbonnier et al. (2010), p. 123.
- ^ Vannier et al. (2006), p. 210.
- ^ Charbonnier et al. (2010), p.119.
- ^ Vannier et al. (2006), p. 210.
- ^ Charbonnier et al. (2010), p. 121.
- ^ Michetti et al., pp. 68-69.
- ^ Lange et al. (2001).
- ^ Hady et al. (2024), capitolo: Sahel Aalma Lagerstätte.
- ^ Vannier et al. (2006), pp. 212-213.
- ^ Van Roy et al. (2021), p. 21.
- ^ Charbonnier et al. (2010), p. 209.
Bibliografia
- (EN) Sylvain Charbonnier, Jean Vannier, Pierre Hantzpergue e Christian Gaillard, Ecological significance of the arthropod fauna from the Jurassic (Callovian) La Voulte Lagerstätte, in Acta Palaeontologica Polonica, vol. 55, n. 1, 2010, pp. 111-132.
- George Hady, Mohamad Bazzi, tamara El Hossny, Nida Ashraf, Pierre Abi Saad e Thomas Clements, The famous fish beds of Lebanon: the Upper Cretaceous Lagerstätten of Haqel, Hjoula, Nammoura, and Sahel Aalma, Journal of the Geological Society, 2024.
- (EN) Carolin Haug, Derek E.G. Briggs, Donald G. Mikulic, Joanne Kluessendorf e Joachim T. Haug, The implications of a Silurian and other thylacocephalan crustaceans for the functional morphology and systematic affinities of the group, in Evolutionary Biology, vol. 14, n. 159, London (UK), 2014, pp. 1-15.
- (EN) Masayuki Ehiro, Osamu Sasaki, Harumasa Kano, Jun Nemoto e Hisaioshi Kato, Thylacocephala (Arthropoda) from the Lower Triassic of the South Kitakami Belt, Northeast Japan, in Paleontological Research, vol. 19, n. 4, 2015, pp. 269-282.
- (EN) Melina Jobbins, Carolin Haug e Christian Klug, First African thylacocephalans from the Famennian of Morocco and their role in Late Devonian food webs, in Scientific reports, vol. 10, n. 5129, 2020, pp. 1-8.
- (EN) Sven Lange, Cees H. Hof, Frederick R. Schram e Fedor E. Steeman, New genus and species from the Cretaceous of Lebanon links the Thylacocephala to the Crustacea, in Palaeontology, vol. 44, n. 5, 2001, pp. 905-912.
- (EN) Thomas Laville, Marie-Béatrice Forel e FSylvain Charbonnier, Re-appraisal of thylacocephalans (Euarthropoda, Thylacocephala) from the Jurassic La Voulte-sur-Rhône Lagerstätte, in European Journal of Taxonomy, vol. 898, 2023, pp. 1-61.
- A.M. Michetti, F. Livio, F.A. Pasquarè, L. Vezzoli, A. Bini, D. Bernoulli e D. Sciunnach, Note illustrative della carta geologica d'Italia - foglio 075 - Como (PDF), Servizio Geologico D'Italia.
- (EN) Stephen Pates, Rudy Lerosey-Aubril, Allison C. Daley, Carlo Kier, Enrico Bonino e Javier Ortega-Hernández, The diverse radiodont fauna from the Marjum Formation of Utah, USA (Cambrian: Drumian), in PeerJ, vol. 9, 2021, pp. 1-43.
- Giovanni Pinna, Paolo Arduini, Carlo Pesarini e Giorgio Teruzzi, Thylacocephala: una nuova classe di crostacei fossili (PDF), in Atti Soc. ital. Sci. nat. Museo civ. Stor. nat. Milano, vol. 123, n. 4, Milano, 1982, pp. 469-482.
- Giovanni Pinna, Exceptional Preservation in the Jurassic of Osteno, in Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, vol. 311, n. 1148, London (UK), 1985, pp. 171-180.
- (EN) Giovanni Pinna, Paolo Arduini, Carlo Pesarini e Giorgio Teruzzi, Some controversial aspects of the morphology and anatomy of Ostenocaris cypriformis (Crustacea, Thylacocephala), in Transactions of the Royal Society of Edinburgh, vol. 76, Edimburgh, 1985, pp. 373–379.
- (EN) Frederick R. Schram, On Mazon Creek Thylacocephala, in Proceedings of the San Diego Society of Natural History, vol. 3, London (UK), 1990, pp. 1–16.
- (EN) Jean Vannier, Chen J.-Y., Huang D.-Y., Sylvain Charbonnier e Wang X.-Q., The Early Cambrian origin of thylacocephalan arthropods, in Acta Palaeontologica Polonica, vol. 51, n. 2, 2006, pp. 201-214.
- (EN) Peter Van Roy, Štěpán Rak, Petr Budil e Oldřich Fatka, Upper Ordovician Thylacocephala (Euarthropoda, Eucrustacea) from Bohemia indicate early ecological differentiation, in Papers in Palaeontology, vol. 7, n. 3, 2021, pp. 1727-1751.
- (EN) H. Zeng, F. Zhao, Z. Yin e M. Zhou, Morphology of diverse radiodontan head sclerites from the early Cambrian Chengjiang Lagerstätte, south-west China, in Journal of Systematic Palaeontology, vol. 16, n. 1, 2018, pp. 1-37.
Altri progetti
Altri progetti
- Wikimedia Commons
 Wikimedia Commons contiene immagini o altri file su Thylacocephala
Wikimedia Commons contiene immagini o altri file su Thylacocephala









